Sensory neurons perceive the environment through specific receptors often enriched within cilia. They communicate these informations to complex neuronal circuits for decision-making. Beside the classical neurotransmission within the synaptic cleft, volume transmission by neuropeptides, and electrical transmission by gap junctions complexify the connectivity pattern of the brain. We explore the underlying cell biology and how this biology can change with age and sex.
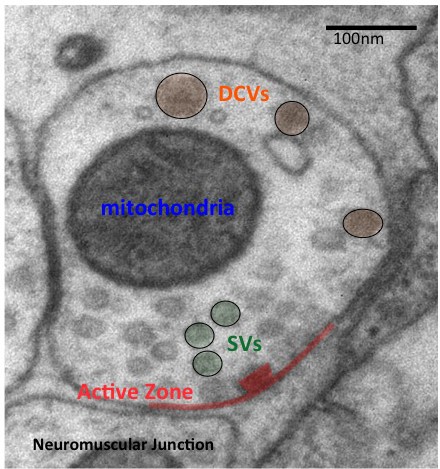
Neurosecretion: How neuropeptides enter the Dense Core Vesicles (DCV)?, how DCV traffic and release are modulated?, how neuropeptides affect nervous system function ? By forward genetics or candidate gene approaches and by behavioral and microscopy readouts, we determined genes involved in neuropeptide release. We identified conserved pathways involved in DCV biogenesis, traffic and secretion in neurons. We showed neuropeptides contribute to cross-talks between sensory neurons within and between circuits.
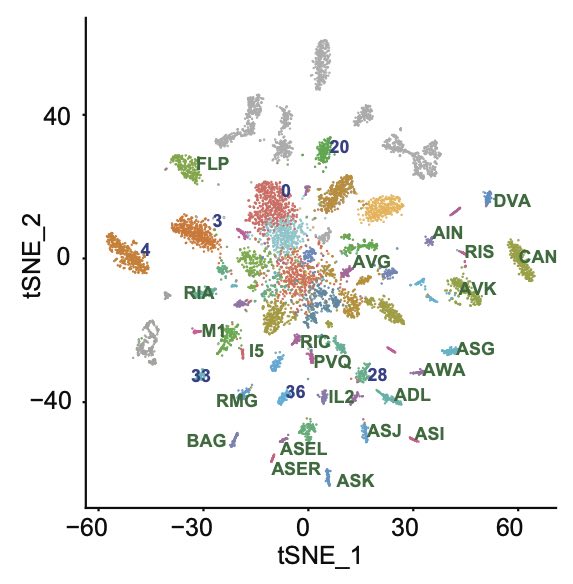
Neuronal atlas: What is the molecular diversity of the C. elegans nervous system?, how does it change over age and sex?, how molecular dimorphisms generate sex-specific behaviour? We generated a molecular atlas of C. elegans, we provided many new cell-specific promoters, we identified sexual dimorphisms within sex-shared neurons.
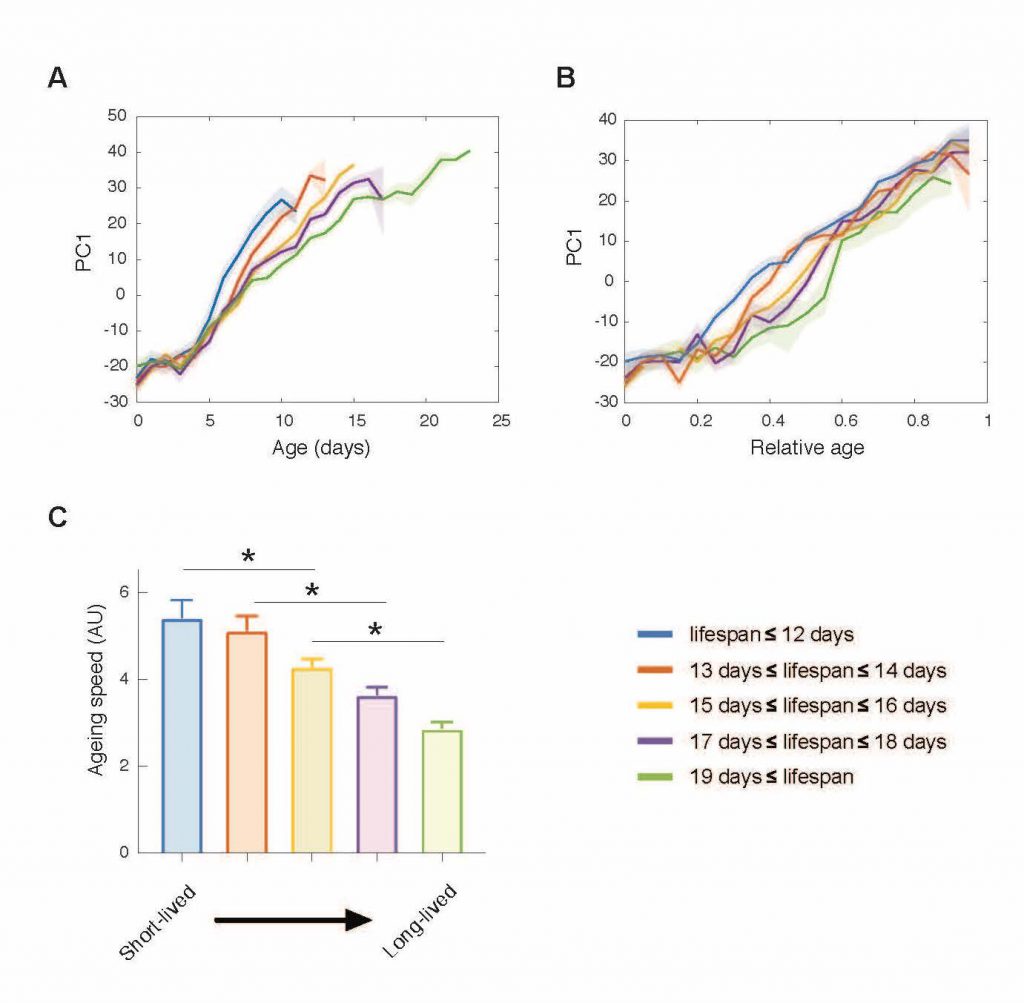
Neuromuscular ageing: We can predict the age of an individual with high accuracy, just based on multiples features of its locomotion. How such a stereotypical behavioural progression is generated by the nervous system? We study gene expresssin change and neuromuscular junction ageing to understand this.
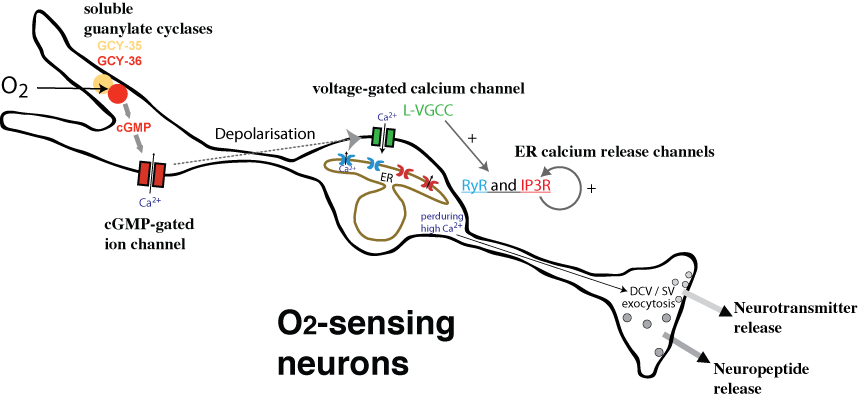
Gaseous sensing: Atmospheric O2 is aversive for C. elegans. During my post-doc, we identified neuroglobin and guanylate cyclases, as necessary sensors for high O2. The O2 sensory neurons are tonic sensors showing little or no adaptation to the cues. Via the release of neuropeptides, they regulate avoidance behaviour and metabolism. More recently we explored the acute H2S-avoidance.
Extracellular vesicles (EVs): How EVs biogenesis and release occur? what are their biological function ? We discovered that most ciliated neurons release EVs from their cilia ending by outward budding of cilia plasma membrane. Ciliary EVs release contributes to maintain cilia composition and sensory function. They may also carry bioactive material for the communication between cells and between individual worms.
